|
|
|
| Molecular and epigenetic pathogenesis of germ cell tumors |
Melanie R. Müllera,Margaretha A.Skowrona,Peter Albersb,Daniel Nettersheima,*( ) )
|
a Department of Urology, Urological Research Lab, Translational UroOncology, University Hospital Düsseldorf, Düsseldorf, Germany
b Department of Urology, University Hospital Düsseldorf, Düsseldorf, Germany |
|
|
|
|
Abstract The development of germ cell tumors (GCTs) is a unique pathogenesis occurring at an early developmental stage during specification, migration or colonization of primordial germ cells (PGCs) in the genital ridge. Since driver mutations could not be identified so far, the involvement of the epigenetic machinery during the pathogenesis seems to play a crucial role. Currently, it is investigated whether epigenetic modifications occurring between the omnipotent two-cell stage and the pluripotent implanting PGCs might result in disturbances eventually leading to GCTs. Although progress in understanding epigenetic mechanisms during PGC development is ongoing, little is known about the complete picture of its involvement during GCT development and eventual classification into clinical subtypes. This review will shed light into the current knowledge of the complex epigenetic and molecular contribution during pathogenesis of GCTs by emphasizing on early developmental stages until arrival of late PGCs in the gonads. We questioned how misguided migrating and/or colonizing PGCs develop to either type I or type II GCTs. Additionally, we asked how pluripotency can be regulated during PGC development and which epigenetic changes contribute to GCT pathogenesis. We propose that SOX2 and SOX17 determine either embryonic stem cell-like (embryonal carcinoma) or PGC-like cell fate (seminoma). Finally, we suggest that factors secreted by the microenvironment, i.e. BMPs and BMP inhibiting molecules, dictate the fate decision of germ cell neoplasia in situ (into seminoma and embryonal carcinoma) and seminomas (into embryonal carcinoma or extraembryonic lineage), indicating an important role of the microenvironment on GCT plasticity.
|
|
Received: 01 November 2019
Available online: 30 May 2020
|
|
Corresponding Authors:
Daniel Nettersheim
E-mail: Daniel.Nettersheim@med.uni-duesseldorf.de
|
|
|
|
|
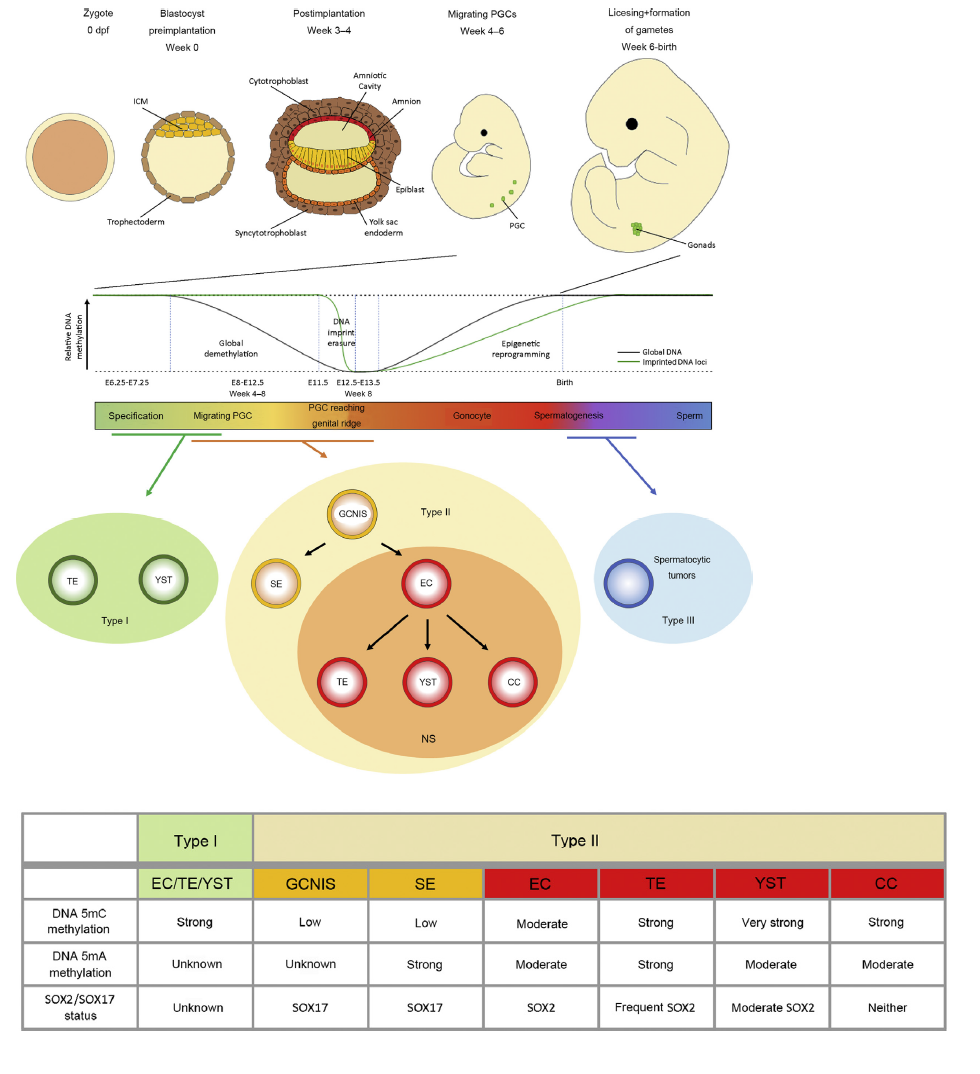
Development of GCs and pathogenesis of TGCTs. Upper panel: Development of the human embryo from the zygote (0 day post fertilization [dpf]) until Week 6 post fertilization when the PGC reach the gonads. Lower panel: DNA methylation events during PGC migration and arrival at the genital ridge and where different types pf TGCTs are thought to originate from. The table summarizes molecular and epigenetic findings from this review. Data from Refs. [1,48,49,52,58,74]. PGC, primordial germ cell; ICM, inner cell mass; SE, seminoma; EC, embryonal carcinoma; CC, choriocarcinoma; TE, teratoma; YST, yolk sac tumor; NS, non-seminoma; GCNIS, germ cell neoplasia in situ; GC, germ cell; TGCTs, testicular germ cell tumors.
|

| Chapter | References | | Comparability of murine and human (primordial) GC development | | The Phenotype differs: Egg cylinder in mice and bilaminar disc in human | [4,19] | | Commonly expressed genes between the species: PRDM1/Prdm1, TFAP2C/Tfap2c, NANOS3/Nanos3, DND1/Dnd1, DDX4/Ddx4, DAZL/Dazl, OCT4/Oct4, NANOG/Nanog | [19] | | Mouse specific genes: Prdm14, Sox2 | [21,22] | | Human specific genes: SOX17, SOX15 | [21,24] | | Development of type I GCTs | | PGC specification depends on WNT and BMP pathways | [27,28] | | The surrounding tissue and extracellular matrix might influence BMP and WNT signaling pathways | this review | | Misrouting of PGCs and failures in downregulating the pluripotency program could lead to type I GCTs | [7] | | Development of type I GCT from PGCs in a short time-frame | [7] | | The testicular dysgenesis syndrome and formation of GCNIS | | TDS results from disturbed hormonal microenvironmental factors during fetal development | [31,32] | | TDS is related to TGCT development | [33] | | Impaired Sertoli and Leydig cell function are not the only triggers leading to GCNIS formation | [33] | | The PGC gene expression program in type II GCTs | | TFAP2C expression is associated with a PGC-like cell fate | [39] | | TFAP2C in GCTs allows maintenance of a latent pluripotent state | this review | | DAZL, DDX4, MAEL and TDRD12 are gonad-specifically activated in PGCs upon arrival at the genital ridge | [40] | | DAZL deficiency leads to an increase in TE formation | [40] | | SOX2 and SOX17 regulating GCT fate | | SOX2 and SOX17 share a common set of overlapping target genes, such as NANOG, OTX2, PIM1/2, PRDM14, DPP4, TDGF1, LIN28A, and TRIM71 | [45] | | Epigenetic re-arrangements might contribute to SOX17 target gene accessibility including GC-fate and pluripotency genes in PGCs | this review | | DNA-Methylation in GCTs | | Compared to SE and GCNIS, NS shows high de novo DNA methylation levels (high expression of DNMT3A and DNMT3L) | [[47], [48], [49]] | | Active 5mC demethylation in TGCTs | [49] | | The influence of microenvironment on TGCT pathogenesis | | Important interactions between SDF1/CXCL12 and CXCR4 as well as functioning Msx genes are involved in a correct migration during PGC development | [[62], [63], [64], [65],67,69] | | A pro-inflammatory micromilieu (IL-1β, IL-6, TNF-α, CCL5, SDF-1/CXCL12, CXCL-13) might favor TGCT development | [68] | | FOXA2 is identified as a key factor of differentiation of SE to NS (not EC) | [44,73] | | Microenvironmental components and BMP-inhibiting factors could directly differentiate SE into EC | [22,44] | | Colonization of the genital ridge is influenced by factors secreted by surrounding Sertoli-, Leydig-, and immune cells, such as chemokines and cytokines | this review |
|
|
Main findings of this review.
|
|
|
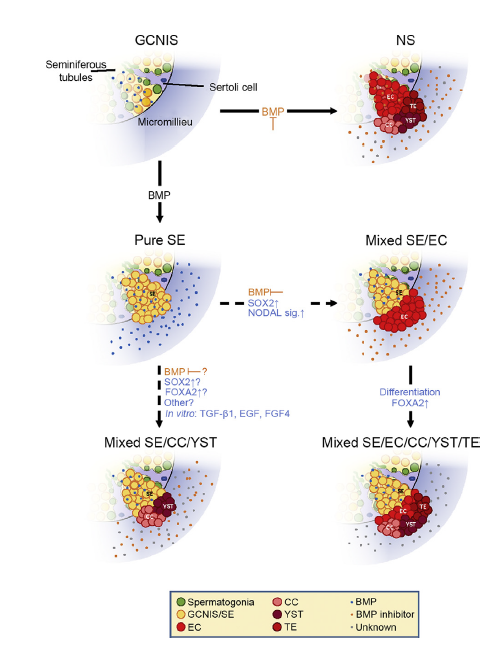
Plasticity of SE cells—Transformation into EC and CC/YST. Depending on BMP activity GCNIS either develop into SE (BMP active) or EC (BMP inhibited) which further differentiates into TE, CC and YST. Additionally, to the commonly accepted TGCT developmental theory (Fig. 1), in vitro experiments indicated reprogramming of SE to EC [22] as well as directly into extraembryonal tumors without EC intermediate (SOX2) [44,75]. SE to EC transformation is initiated in vitro by BMP inhibition through the microenvironment. SOX2 upregulation and establishment of NODAL signaling give rise to mixed SE/EC. These can further differentiate into mixed SE/NS, eventually leading to mixed tumors with SE components while also containing extraembryonal proportions. Direct SE differentiation into extraembryonal-like tissue has been demonstrated in vitro and in vivo although the factors involved in the latter still remain unknown and have yet to be determined. GCNIS, germ cell neoplasia in situ; EC, embryonal carcinoma; SE, seminoma; NS, non-seminoma; CC, choriocarcinoma; TE, teratoma; YST, yolk sac tumor; NS, non-seminoma; GC, germ cell; TGCT, testicular germ cell tumor.
|
| [1] |
J.W. Oosterhuis, L.H.J. Looijenga, Human germ cell tumours from a developmental perspective, Nat Rev Canc, 19(2019), pp.522-537
doi: 10.1038/s41568-019-0178-9
|
| [2] |
R.D. Palmer, N.A. Foster, S.L. Vowler, I. Roberts, C.M. Thornton, J.P. Hale, et al. Malignant germ cell tumours of childhood: new associations of genomic imbalance, Br J Canc, 96(2007), pp. 667-676
doi: 10.1038/sj.bjc.6603602
|
| [3] |
A. Fonseca, A.L. Frazier, F. Shaikh, Germ cell tumors in adolescents and young adults, J Oncol Pract, 15(2019), pp. 433-441
doi: 10.1200/JOP.19.00190
|
| [4] |
E.G. Baraban, K. Cooper, Pathogenesis of testicular germ cell neoplasia: a conceptual approach, Adv Anat Pathol, 26(2019), pp. 241-245
doi: 10.1097/PAP.0000000000000233
pmid: 30950848
|
| [5] |
H. Hernandez-Vargas, N. Sincic, M. Ouzounova, Z. Herceg, Epigenetic signatures in stem cells and cancer stem cells, Epigenomics, 1(2009), pp. 261-280
doi: 10.2217/epi.09.19
pmid: 22122702
|
| [6] |
D.M. Berney, L.H.J. Looijenga, M. Idrees, J.W. Oosterhuis, E. Rajpert-De Meyts, T.M. Ulbright, et al. Germ cell neoplasia in situ (GCNIS): evolution of the current nomenclature for testicular pre-invasive germ cell malignancy, Histopathology, 69(2016), pp. 7-10
doi: 10.1111/his.2016.69.issue-1
|
| [7] |
J.W. Oosterhuis, L.H.J. Looijenga, Testicular germ-cell tumours in a broader perspective, Nat Rev Canc, 5(2005), pp. 210-222
doi: 10.1038/nrc1568
|
| [8] |
K. Hayashi, SMC de Sousa Lopes, M.A. Surani, Germ cell specification in mice, Science, 316(2007), pp. 394-396
doi: 10.1126/science.1137545
|
| [9] |
D.G. Kristensen, N.E. Skakkeb?k, E. Rajpert-De Meyts, K. Almstrup, Epigenetic features of testicular germ cell tumours in relation to epigenetic characteristics of foetal germ cells, Int J Dev Biol, 57(2013), pp. 309-317
doi: 10.1387/ijdb.130142ka
|
| [10] |
P.K. Suresh, Mechanisms of pluripotency and epigenetic reprogramming in primordial germ cells: lessons for the conversion of other cell types into the stem cell lineage, Turk J Biol, 39(2015), pp. 187-193
doi: 10.3906/biy-1407-16
|
| [11] |
K. Kurimoto, Y. Yabuta, Y. Ohinata, M. Shigeta, K. Yamanaka, M. Saitou, Complex genome-wide transcription dynamics orchestrated by Blimp1 for the specification of the germ cell lineage in mice, Genes Dev, 22(2008), pp. 1617-1635
doi: 10.1101/gad.1649908
|
| [12] |
K. Mochizuki, Y. Tando, T. Sekinaka, K. Otsuka, Y. Hayashi, H. Kobayashi, et al. SETDB1 is essential for mouse primordial germ cell fate determination by ensuring BMP signaling SETDB1 is essential for mouse primordial germ cell fate determination by ensuring BMP signaling, Development, 145 (2018), dev164160, 10.1242/dev.164160.
|
| [13] |
J.K. Killian, L.C.J. Dorssers, B. Trabert, A.J.M. Gillis, M.B. Cook, Y. Wang, et al. Imprints and DPPA3 are bypassed during pluripotency- and differentiation-coupled methylation reprogramming in testicular germ cell tumors, Genome Res, 26(2016), pp. 1490-1504
doi: 10.1101/gr.201293.115
|
| [14] |
B.E. Richardson, R. Lehmann, Mechanisms guiding primordial germ cell migration: strategies from different organisms, Nat Rev Mol Cell Biol, 11(2010), pp. 37-49
doi: 10.1038/nrm2815
|
| [15] |
D. Eckert, K. Biermann, D. Nettersheim, A.J.M. Gillis, K. Steger, H.-M. J?ck, et al. Expression of BLIMP1/PRMT5 and concurrent histone H2A/H4 arginine 3 dimethylation in fetal germ cells, CIS/IGCNU and germ cell tumors,BMC Dev Biol, 8(2008), p. 106
|
| [16] |
M de Felici, Nuclear reprogramming in mouse primordial germ cells: epigenetic contribution, Stem Cell Int, 2011 (2011), p. 425863, 10.4061/2011/425863.
|
| [17] |
H. Sasaki, Y. Matsui, Epigenetic events in mammalian germ-cell development: reprogramming and beyond, Nat Rev Genet, 9(2008), pp. 129-140
doi: 10.1038/nrg2295
|
| [18] |
Y.C. Hu, P.K. Nicholls, Y.Q. Soh, J.R. Daniele, J.P. Junker, A. van Oudenaarden, et al. Licensing of primordial germ cells for gametogenesis depends on genital ridge signaling, PLoS Genet, 11 (2015), Article e1005019, 10.1371/journal.pgen.1005019.
|
| [19] |
T. Kobayashi, M.A. Surani, On the origin of the human germline, Development, 145(2018), p. dev150433, 10.1242/dev.150433.
|
| [20] |
I. Aksoy, R. Jauch, J. Chen, M. Dyla, U. Divakar, G.K. Bogu, et al. Oct4 switches partnering from Sox2 to Sox17 to reinterpret the enhancer code and specify endoderm, EMBO J, 32(2013), pp. 938-953
doi: 10.1038/emboj.2013.31
|
| [21] |
N. Irie, L. Weinberger, W.W.C. Tang, T. Kobayashi, S. Viukov, Y.S. Manor, et al. SOX17 is a critical specifier of human primordial germ cell fate, Cell, 160(2015), pp. 253-268
doi: 10.1016/j.cell.2014.12.013
|
| [22] |
D. Nettersheim, S. Jostes, R. Sharma, S. Schneider, A. Hofmann, H.J. Ferreira, et al. BMP inhibition in seminomas initiates acquisition of pluripotency via NODAL signaling resulting in reprogramming to an embryonal carcinoma, Article e1005415, 11(2015), Article e1005415, 10.1371/journal.pgen.1005415.
|
| [23] |
A. Sybirna, W.W.C. Tang, M. Pierson Smela, S. Dietmann, W.H. Gruhn, R. Brosh, et al. A critical role of PRDM14 in human primordial germ cell fate revealed by inducible degrons, Nat Commun, 11 (2020), 10.1038/s41467-020-15042-0.
|
| [24] |
M. Pierson Smela, A. Sybirna, F.C.K. Wong, M.A. Surani, Testing the role of SOX15 in human primordial germ cell fate, Wellcome Open Res, 4 (2019), p. 122, 10.12688/wellcomeopenres.15381.2.
|
| [25] |
J.L. Pierce, A.L. Frazier, J.F. Amatruda, Pediatric germ cell tumors: a developmental perspective, Adv Urol, 2018 (2018), p. 9059382, 10.1155/2018/9059382.
|
| [26] |
A. Murrell, Genomic imprinting and cancer: from primordial germ cells to somatic cells, Sci World J, 6(2006), pp. 1888-1910
pmid: 17205195
|
| [27] |
J.A. Hackett, Y. Huang, U. Günesdogan, K.A. Gretarsson, T. Kobayashi, M.A. Surani, Tracing the transitions from pluripotency to germ cell fate with CRISPR screening, Nat Commun, 9 (2018), p. 4292, 10.1038/s41467-018-06230-0.
doi: 10.1038/s41467-018-06230-0
pmid: 30327475
|
| [28] |
N. Fustino, D. Rakheja, C.S. Ateek, J.C. Neumann, J.F. Amatruda, Amatruda,Bone morphogenetic protein signalling activity distinguishes histological subsets of paediatric germ cell tumours, Int J Androl, 34 (2011), pp. e218-e233, 10.1111/j.1365-2605.2011.01186.x.
doi: 10.1111/j.1365-2605.2011.01186.x
|
| [29] |
I. Bejarano, A.B. Rodríguez, J.A. Pariente, Apoptosis is a demanding selective tool during the development of fetal male germ cells, Front Cell Dev Biol, 6 (2018), 10.3389/fcell.2018.00065.
|
| [30] |
N.E. Skakkebaek, M. Holm, C. Hoei-Hansen, N. J?rgensen, E. Rajpert-De Meyts, Association between testicular dysgenesis syndrome (TDS) and testicular neoplasia: evidence from 20 adult patients with signs of maldevelopment of the testis, APMIS, 111(2003), pp. 1-11
pmid: 12752226
|
| [31] |
N.E. Skakkebaek, E. Rajpert-De Meyts, K.M. Main, Testicular dysgenesis syndrome: an increasingly common developmental disorder with environmental aspects, Hum Reprod, 16(2001), pp. 972-978
pmid: 11331648
|
| [32] |
S.B. Sonne, D.M. Kristensen, G.W. Novotny, I.A. Olesen, J.E. Nielsen, N.E. Skakkebaek, et al. Testicular dysgenesis syndrome and the origin of carcinoma in situ testis, Int J Androl, 31(2008), pp. 275-287
doi: 10.1111/ija.2008.31.issue-2
|
| [33] |
C.E. Hoei-Hansen, M. Holm, E. Rajpert-De Meyts, N.E. Skakkebaek, Histological evidence of testicular dysgenesis in contralateral biopsies from 218 patients with testicular germ cell cancer, J Pathol, 200(2003), pp. 370-374
pmid: 12845633
|
| [34] |
C. Chen, W. Ouyang, V. Grigura, Q. Zhou, K. Carnes, H. Lim, et al. ERM is required for transcriptional control of the spermatogonial stem cell niche, Nature, 436(2005), pp. 1030-1034
pmid: 16107850
|
| [35] |
M. Szarek, M. Bergmann, L. Konrad, H.-C. Schuppe, S. Kliesch, M.P. Hedger , et al. Activin A target genes are differentially expressed between normal and neoplastic adult human testes: clues to gonocyte fate choice, Andrology, 7(2019), pp. 31-41
doi: 10.1111/andr.12553
pmid: 30315637
|
| [36] |
R.T. Dorsam, J.S. Gutkind, G-protein-coupled receptors and cancer, Nat Rev Canc, 7(2007), pp. 79-94
doi: 10.1038/nrc2069
|
| [37] |
J.C. Young, A. Jaiprakash, S. Mithraprabhu, C. Itman, R. Kitazawa, L.H.J. Looijenga, et al. TCam-2 seminoma cell line exhibits characteristic foetal germ cell responses to TGF-beta ligands, Int J Androl, 34 (2011), pp. e204-e217, 10.1111/j.1365-2605.2011.01170.x.
doi: 10.1111/j.1365-2605.2011.01170.x
|
| [38] |
J. de Jong, H. Stoop, A.J. Gillis, R.J. van Gurp, G.J. van de Geijn, Md Boer, et al. Differential expression of SOX17 and SOX2 in germ cells and stem cells has biological and clinical implications, J Pathol, 215(2008), pp. 21-30
doi: 10.1002/path.2332
pmid: 18348160
|
| [39] |
S. Weber, D. Eckert, D. Nettersheim, A.J. Gillis, S. Sch?fer, P. Kuckenberg, et al. Critical function of AP-2 gamma/TCFAP2C in mouse embryonic germ cell maintenance, Biol Reprod, 82(2010), pp. 214-223
doi: 10.1095/biolreprod.109.078717
|
| [40] |
P.K. Nicholls, H. Schorle, S. Naqvi, Y.C. Hu, Y. Fan, M.A. Carmell, et al. Mammalian germ cells are determined after PGC colonization of the nascent gonad, Proc Natl Acad Sci U S A, 116(2019), pp. 25677-25687
doi: 10.1073/pnas.1910733116
pmid: WOS:000503281500050
|
| [41] |
K. Takahashi, S. Yamanaka, Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors, Cell, 126(2006), pp. 663-676
doi: 10.1016/j.cell.2006.07.024
|
| [42] |
D. Nettersheim, S. Jostes, S. Schneider, H. Schorle, Elucidating human male germ cell development by studying germ cell cancer, Reproduction, 152 (2016), pp. R101-R113, 10.1530/REP-16-0114.
doi: 10.1530/REP-16-0114
|
| [43] |
H.G. Leitch, A. Smith, The mammalian germline as a pluripotency cycle, Development, 140(2013), pp. 2495-2501
doi: 10.1242/dev.091603
pmid: 23715543
|
| [44] |
D. Nettersheim, A. Heimsoeth, S. Jostes, S. Schneider, M. Fellermeyer, A. Hofmann, et al. SOX2 is essential for in vivo reprogramming of seminoma-like TCam-2 cells to an embryonal carcinoma-like fate, Oncotarget, 7(2016), pp. 47095-47110
doi: 10.18632/oncotarget.9903
pmid: 27283990
|
| [45] |
S.V. Jostes, M. Fellermeyer, L. Arévalo, G.E. Merges, G. Kristiansen, D. Nettersheim, et al. Unique and redundant roles of SOX2 and SOX17 in regulating the germ cell tumour fate, Int J Canc, 146(2019), pp. 1592-1605
doi: 10.1002/ijc.v146.6
|
| [46] |
Y. Kojima, K. Sasaki, S. Yokobayashi, Y. Sakai, T. Nakamura, Y. Yabuta, et al. Evolutionarily distinctive transcriptional and signaling programs drive human germ cell lineage specification from pluripotent stem cells, Cell Stem Cell, 21 (2017), pp. 517-532.e5, 10.1016/j.stem.2017.09.005.
|
| [47] |
J. Lobo, R. Henrique, C. Jerónimo, The role of DNA/histone modifying enzymes and chromatin remodeling complexes in testicular germ cell tumors, Cancers (Basel), 11 (2018), p. E6, 10.3390/cancers11010006.
|
| [48] |
G.J. Netto, Y. Nakai, M. Nakayama, S. Jadallah, A. Toubaji, N. Nonomura, et al. Global DNA hypomethylation in intratubular germ cell neoplasia and seminoma, but not in nonseminomatous male germ cell tumors, Mod Pathol, 21(2008), pp. 1337-1344
doi: 10.1038/modpathol.2008.127
|
| [49] |
D. Nettersheim, L.C. Heukamp, F. Fronhoffs, M.J. Grewe, N. Haas, A. Waha, et al. Analysis of TET expression/activity and 5mC oxidation during normal and malignant germ cell development, PloS One, 8 (2013), Article e82881, 10.1371/journal.pone.0082881.
|
| [50] |
D.J. Smiraglia, J. Szymanska, S.M. Kraggerud, R.A. Lothe, P. Peltom?ki, C. Plass, Distinct epigenetic phenotypes in seminomatous and nonseminomatous testicular germ cell tumors, Oncogene, 21(2002), pp. 3909-3916
pmid: 12032829
|
| [51] |
G.E. Lind, R.I. Skotheim, R.A. Lothe, The epigenome of testicular germ cell tumors, APMIS, 115(2007), pp. 1147-1160
doi: 10.1111/apm.2007.115.issue-10
|
| [52] |
A.L. Costa, C. Moreira-Barbosa, J. Lobo, B. Vilela-Salgueiro, M. Cantante, R. Guimar?es, et al. DNA methylation profiling as a tool for testicular germ cell tumors subtyping, Epigenomics, 10(2018), pp. 1511-1523
doi: 10.2217/epi-2018-0034
pmid: 30418048
|
| [53] |
J. Lobo, A.J.M. Gillis, C. Jerónimo, R. Henrique, L.H.J. Looijenga, Human germ cell tumors are developmental cancers: impact of epigenetics on pathobiology and clinic, Int J Mol Sci, 20 (2019), p. E258, 10.3390/ijms20020258.
|
| [54] |
P. Ji, X. Wang, N. Xie, Y. Li, N6-Methyladenosine in RNA and DNA: an epitranscriptomic and epigenetic player implicated in determination of stem cell fate, Stem Cell Int, 2018 (2018), p. 3256524, 10.1155/2018/3256524.
|
| [55] |
D. Nettersheim, D. Berger, S. Jostes, G. Kristiansen, G. Lochnit, H. Schorle, N6-Methyladenosine detected in RNA of testicular germ cell tumors is controlled by METTL3, ALKBH5, YTHDC1/F1/F2, and HNRNPC as writers, erasers, and readers, Andrology, 7(2019), pp. 498-506
doi: 10.1111/andr.12612
pmid: 30903744
|
| [56] |
P.J. Batista, B. Molinie, J. Wang, K. Qu, J. Zhang, L. Li, et al. m(6)A RNA modification controls cell fate transition in mammalian embryonic stem cells, Cell Stem Cell, 15(2014), pp. 707-719
doi: 10.1016/j.stem.2014.09.019
pmid: 25456834
|
| [57] |
S. Schwartz, M.R. Mumbach, M. Jovanovic, T. Wang, K. Maciag, G.G. Bushkin, et al. Perturbation of m6A writers reveals two distinct classes of mRNA methylation at internal and 5’ sites, Cell Rep, 8(2014), pp. 284-296
doi: 10.1016/j.celrep.2014.05.048
pmid: 24981863
|
| [58] |
J. Lobo, A.L. Costa, M. Cantante, R. Guimaraes, P. Lopes, L. Antunes, , et al. m6A RNA modification and its writer/reader VIRMA/YTHDF3 in testicular germ cell tumors: a role in seminoma phenotype maintenance, J Transl Med, 17 (2019), 10.1186/s12967-019-1837-z.
|
| [59] |
Y. Wang, Y. Li, J.I. Toth, M.D. Petroski, Z. Zhang, J.C. Zhao, N6-methyladenosine modification destabilizes developmental regulators in embryonic stem cells, Nat Cell Biol, 16(2014), pp. 191-198
doi: 10.1038/ncb2902
|
| [60] |
M. Oliveros-Etter, Z. Li, K. Nee, L. Hosohama, J. Hargan-Calvopina, S.A. Lee, et al. PGC reversion to pluripotency involves erasure of DNA methylation from imprinting control centers followed by locus-specific re-methylation, Stem Cell Reports, 5(2015), pp. 337-349
doi: 10.1016/j.stemcr.2015.07.006
pmid: 26278040
|
| [61] |
K. Hayashi, H. Ohta, K. Kurimoto, S. Aramaki, M. Saitou, Reconstitution of the mouse germ cell specification pathway in culture by pluripotent stem cells, Cell, 146(2011), pp. 519-532
doi: 10.1016/j.cell.2011.06.052
|
| [62] |
T. Ara, Y. Nakamura, T. Egawa, T. Sugiyama, K. Abe, T. Kishimoto, et al. Impaired colonization of the gonads by primordial germ cells in mice lacking a chemokine, stromal cell-derived factor-1 (SDF-1), Proc Natl Acad Sci U S A, 100(2003), pp. 5319-5323
doi: 10.1073/pnas.0730719100
|
| [63] |
M. Doitsidou, M. Reichman-Fried, J. Stebler, M. K?prunner, J. D?rries, D. Meyer, et al. Guidance of primordial germ cell migration by the chemokine SDF-1, Cell, 111(2002), pp. 647-659
pmid: 12464177
|
| [64] |
K.A. Molyneaux, H. Zinszner, P.S. Kunwar, K. Schaible, J. Stebler, M.J. Sunshine, et al. The chemokine SDF1/CXCL12 and its receptor CXCR4 regulate mouse germ cell migration and survival, Development, 130(2003), pp. 4279-4286
pmid: 12900445
|
| [65] |
H. Li, R. Liang, Y. Lu, M. Wang, Z. Li, RTN3 regulates the expression level of chemokine receptor CXCR4 and is required for migration of primordial germ cells, Int J Mol Sci, 17 (2016), p. 382, 10.339.0/ijms17040382
doi: 10.3390/ijms17040382
|
| [66] |
D.C. Gilbert, I. Chandler, A. McIntyre, N.C. Goddard, R. Gabe, R.A. Huddart, et al. Clinical and biological significance of CXCL12 and CXCR4 expression in adult testes and germ cell tumours of adults and adolescents, J Pathol, 217(2009), pp. 94-102
doi: 10.1002/path.v217:1
|
| [67] |
J. Stebler, D. Spieler, K. Slanchev, K.A. Molyneaux, U. Richter, V. Cojocaru, et al. Primordial germ cell migration in the chick and mouse embryo: the role of the chemokine SDF-1/CXCL12, Dev Biol, 272(2004), pp. 351-361
pmid: 15282153
|
| [68] |
T. Lei, S. Moos, J. Klug, F. Aslani, S. Bhushan, E. Wahle, et al. Galectin-1 enhances TNFα-induced inflammatory responses in Sertoli cells through activation of MAPK signalling, Sci Rep, 8 (2018), p. 3741, 10.1038/s41598-018-22135-w.
doi: 10.1038/s41598-018-22135-w
|
| [69] |
B. Klein, T. Haggeney, D. Fietz, S. Indumathy, K.L. Loveland, M. Hedger, et al. Specific immune cell and cytokine characteristics of human testicular germ cell neoplasia, Hum Reprod, 31(2016), pp. 2192-2202
doi: 10.1093/humrep/dew211
|
| [70] |
A. Alvarez-Buylla, H. Merchant-Larios, Mouse primordial germ cells use fibronectin as a substrate for migration, Exp Cell Res, 165(1986), pp. 362-368
pmid: 3720854
|
| [71] |
X. Sun, C.B. Park, W. Deng, S.S. Potter, S.K. Dey, Uterine inactivation of muscle segment homeobox (Msx) genes alters epithelial cell junction proteins during embryo implantation, Faseb J, 30(2016), pp. 1425-1435
doi: 10.1096/fsb2.v30.4
|
| [72] |
C.M. Spiller, C.-W. Feng, A. Jackson, A.J.M. Gillis, A.D. Rolland, L.H.J. Looijenga, et al. Endogenous Nodal signaling regulates germ cell potency during mammalian testis development, Development, 139(2012), pp. 4123-4132
doi: 10.1242/dev.083006
pmid: 23034635
|
| [73] |
D. Nettersheim, S. Vadder, S. Jostes, A. Heimsoeth, H. Schorle, TCam-2 cells deficient for SOX2 and FOXA2 are blocked in differentiation and maintain a seminoma-like cell fate in vivo, Cancers (Basel), 11 (2019), p. E728, 10.3390/cancers11050728.
|
| [74] |
D. Nonaka, Differential expression of SOX2 and SOX17 in testicular germ cell tumors, Am J Clin Pathol, 131(2009), pp. 731-736
doi: 10.1309/AJCP7MNCNBCRN8NO
|
| [75] |
D. Nettersheim, A.J. Gillis, L.H. Looijenga, H. Schorle, TGF-β1, EGF and FGF4 synergistically induce differentiation of the seminoma cell line TCam-2 into a cell type resembling mixed non-seminoma, Int J Androl, 34 (2011), pp. e189-e203, 10.1111/j.1365-2605.2011.01172.x.
doi: 10.1111/j.1365-2605.2011.01172.x
|
| [1] |
Christian Winter,Andreas Hiester. Treatment of clinical stage I non-seminoma[J]. Asian Journal of Urology, 2021, 8(2): 161-169. |
| [2] |
Takeshi Hirata, Seung Chol Park, Michelle T. Muldong, Christina N. Wu, Tomonori Yamaguchi, Amy Strasner, Omer Raheem, Hiromi Kumon, Robert L. Sah, Nicholas A. Cacalano, Catriona H. M. Jamieson, Christopher J. Kane, Koichi Masuda, Anna A. Kulidjian, Christina A. M. Jamieson. Specific bone region localization of osteolytic versus osteoblastic lesions in a patient-derived xenograft model of bone metastatic prostate cancer[J]. Asian Journal of Urology, 2016, 3(4): 229-239. |
|
|
|
|